Introduction
Headache is a common concern among children and adolescents, with a lifetime prevalence of 60.1% in pediatric patients in Korea [1]. Primary headaches, including migraines and tension-type headaches, arise from a variety of causes. Environmental and emotional factors, including stress, psychiatric disorders, and adverse childhood experiences, play key roles in triggering these headaches [2].
In 2019, the emergence of the coronavirus disease 2019 (COVID-19) pandemic profoundly impacted human lives. Following the initial outbreak in February 2020, Korean students were prohibited from attending school. Starting in mid-April 2020, online classes were gradually implemented, leading to the eventual shutdown of most in-person instruction [3]. Recent data from Korea regarding the impact of the prolonged COVID-19 pandemic on the mental health of Korean adolescents reveal a troubling link between preventive measures, such as school closures and remote classes, and increased anxiety and depressive symptoms in this population [4].
Scant research has been published examining behavioral and emotional changes in pediatric patients with headaches, from before to after the onset of the COVID-19 pandemic. Consequently, we conducted a comparative analysis to investigate these changes within this demographic. The present study was designed to identify any significant behavioral or emotional changes among pediatric patients with headaches following the onset of the COVID-19 pandemic and to examine the impact of these changes on pediatric headaches.
Materials and Methods
We conducted a retrospective review of data from 162 patients who visited the pediatric headache clinic at Hallym University Kangnam Sacred Heart Hospital (HUKSHH) between March 2018 and August 2021. Patients were categorized into two groups based on the timing of pandemic onset: the before pandemic (BP) group and the after pandemic (AP) group. The BP group included 81 patients who visited the clinic during the pre-pandemic period (March 2018 to August 2019), while the AP group comprised 81 patients who visited after the pandemic had begun (January 2020 to August 2021). The Institutional Review Board of HUKSHH reviewed and approved this study (2022-05-033). Due to the retrospective nature of the research, the requirement for informed consent from the participants was waived.
The study included participants aged 6 to 19 years who were diagnosed with primary headache according to the criteria outlined in the third edition of the International Classification of Headache Disorders [5]. During the baseline survey, we collected biographical data, lifestyle characteristics, general medical histories, and headache-related information, including headache frequency, clinical features, and duration. Imaging studies, including brain magnetic resonance imaging and cervical spine radiography, were also performed.
Behavioral and psychiatric screening tests, such as the Korean Child Behavior Checklist (K-CBCL), the State Anxiety Inventory for Children (SAIC), the Trait Anxiety Inventory for Children (TAIC), and the Children’s Depression Inventory (CDI), were administered. Patients who did not complete one or more of these questionnaires were excluded.
Statistical analyses were performed using SPSS version 28 (IBM Corp., Armonk, NY, USA). The Mann-Whitney U test was utilized to compare headache characteristics and scores on the K-CBCL, SAIC, TAIC, and CDI between the groups. Correlation analysis was employed to examine the relationships among SAIC, TAIC, CDI, and K-CBCL scores. P values of less than 0.05 were considered to indicate statistical significance, and all P values were two-tailed.
Results
The study included 162 patients, with a male-to-female ratio of 63:99 and a mean age of 11.7±3.3 years. These patients were categorized into BP and AP groups (n=81 for each). No significant differences were noted between these groups regarding age, age at headache onset, headache severity, disruption in daily activities due to headache, aggravation of headache by daily activities, associated dizziness, family history of headache, or history of psychiatric consultation. Additionally, the average frequency of headaches was similar between groups, with the BP group experiencing headaches an average of 4.49 times per week and the AP group 4.04 times per week. The mean duration of headache was 6.34 hours for the BP group and 10.07 hours for the AP group; however, this difference was not statistically significant. A higher proportion of patients in the BP group reported disruptions in daily activities due to headaches (24.69%) (Table 1).
Among the 81 patients in the BP group, the characteristics of pain were most commonly described as pulsating (48.2%, n=39), followed by pressing (23.5%, n=19) and cramping (9.9%, n=8). In the AP group (also comprising 81 participants), the primary descriptions were pulsating (55.6%, n=45), pressing (17.3%, n=14), and other (9.9%, n=8). No significant difference was observed between the groups in terms of these pain characteristics (P=0.486).
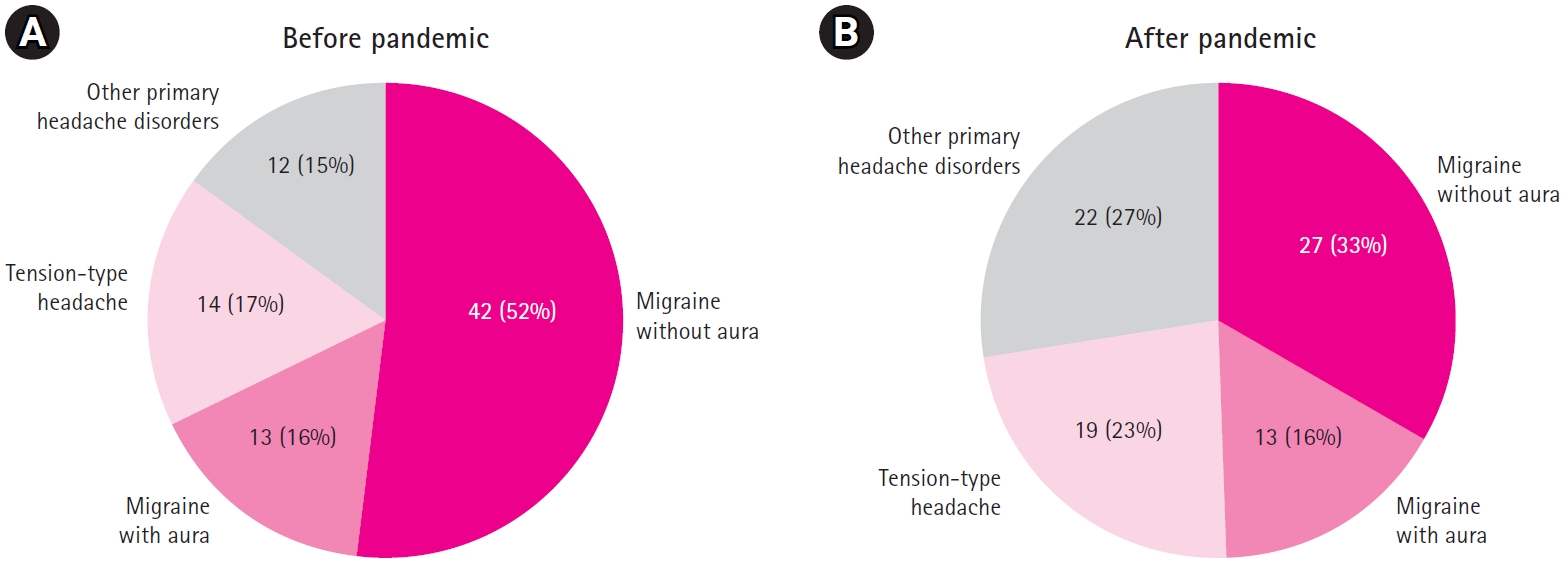
Regarding final diagnosis, the combined rate of migraine without aura and migraine with aura was 68% in the BP group, which exceeded that in the AP group. The AP participants exhibited a higher proportion of diagnoses categorized as primary headache disorders other than migraine or tension-type headaches. Overall, a significant difference in diagnosis was observed between the two groups (P=0.009). However, when patients were further classified into diagnostic subgroups and the K-CBCL, SAIC, TAIC, and CDI values were compared, no statistically significant differences were found (Fig. 1).
After incorporating the K-CBCL results into the comparative analysis, no significant difference was observed between the two groups for any of the detailed scores, including the K-CBCL total problem score (Table 2).
The mean CDI and SAIC scores were significantly higher in the AP group compared to the BP group. For the AP group, chi-square test results indicated an odds ratio of 4.971 for an abnormal CDI score. However, the chi-square test revealed no significant association. Specifically, regarding CDI, three patients in the BP group and 13 in the AP group displayed scores in the abnormal range, indicating a higher proportion of such scores in the AP group. In terms of SAIC, 13 patients in the BP group and 19 in the AP group had values in the abnormal range. Although the average TAIC scores did not differ significantly between groups, the AP group contained more patients with abnormal values (n=19) than the BP group (n=11), as shown in Table 3.
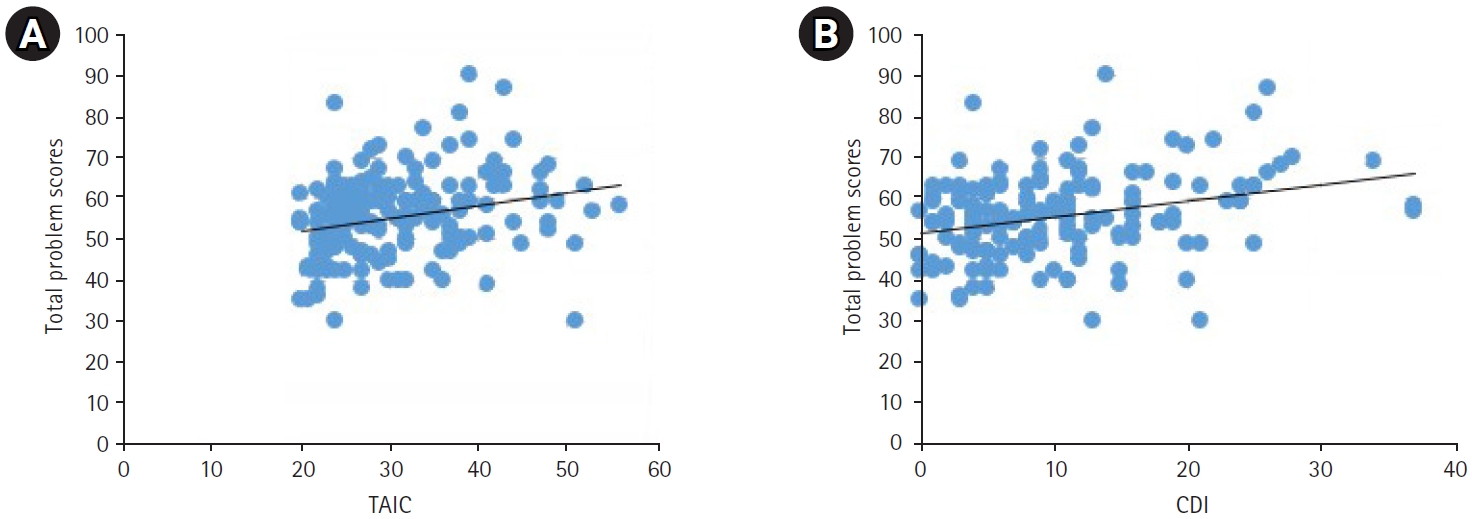
When analyzing the correlation between SAIC, TAIC, CDI, and the total problem score of K-CBCL, it was found that there was no statistically significant correlation between SAIC and the K-CBCL total problem scores. However, a weak positive correlation was observed between the TAIC score (Pearson correlation coefficient, 0.223) and the K-CBCL total problem score (P=0.002). Similarly, the CDI score demonstrated a weak positive correlation (coefficient, 0.297) with the total problem score (P<0.001) (Fig. 2).
Discussion
The COVID-19 pandemic has meaningfully altered the daily lives of children and adolescents. The time spent in classes at school and sessions at private tutoring centers has decreased, whereas smartphone usage has risen. Time spent exercising has declined by 55%, contributing to a 33% increase in student body weight [3]. The pandemic was associated with increased stress levels (60.6%), reduced physical activity (77.2%), moderate-to-severe concerns about COVID-19 (84.3%), and worsened mood and sleep changes [6].
These alterations have also negatively impacted individuals with migraine. Research has consistently shown that environmental and psychological factors, sleep disturbances, and fatigue can trigger primary headaches [7,8]. In the context of COVID-19, 33.8% of patients with migraine reported an increase in the use of acute treatments for migraine attacks, while 18.8% described changes or additions to their preventive medications. Among 325 pediatric patients in Korea, 18.5% experienced a significant worsening of migraine headaches compared to their usual condition [6]. Furthermore, among 607 patients with migraine, 59.6% reported an increase in attack frequency, 16% reported a decrease, and 10.3% transitioned to chronic migraine. The headache severity increased in 64.1% of cases. Sleep disturbances were reported by 78.1% of patients, and symptoms of anxiety and/or depression were observed in 79.5% [9].
In this study, we examined the impact of COVID-19 on pediatric patients with primary headaches. We found no significant differences in headache frequency, intensity, or duration between the BP and AP groups. However, a higher proportion of patients in the BP group reported disruptions in daily activities due to headaches, which may relate to school attendance—an important aspect of daily life for children and adolescents.
Interestingly, the average SAIC and CDI scores were significantly higher in the AP group than in the BP group. While the average TAIC score was also higher in the AP cohort, this difference did not reach statistical significance. These findings suggest that the COVID-19 pandemic has influenced the emotional well-being of pediatric patients with headaches. The distinction between SAIC, which measures “state” anxiety, and TAIC, which measures “trait” anxiety, is particularly noteworthy in considering how the pandemic may influence patients’ emotional states. These results suggest the need to examine the effects of the pandemic not only in terms of headache frequency or intensity but also from an emotional perspective.
In contrast, the results regarding the K-CBCL, which is used to evaluate behavioral problems, revealed no significant differences in the total problem scores or detailed item scales between the two groups. This outcome contradicts the hypothesis that the COVID-19 pandemic had triggered behavioral changes in pediatric and adolescent patients.
However, a statistically significant correlation was found between the total K-CBCL problem scores and both TAIC and CDI scores. Furthermore, a significant increase in CDI scores was observed in the AP group. A study involving 4,702 pediatric participants revealed a significant deterioration in CBCL scores during the COVID-19 pandemic among children whose pre-pandemic scores were within the normal range. However, the same research also noted instances of improvement, particularly among children who had already experienced severe mental health symptoms before the pandemic [10]. In light of these findings, further research should be undertaken with a larger sample and a more detailed analysis of patient groups.
This study has several limitations. First, we did not observe headache patterns in the same patients before and after the onset of COVID-19. Our focus was on distinct patients who sought medical attention during the pre- and post-pandemic periods, and the observed differences may have been influenced by individual variations. Second, healthy controls were not included in the analysis. Instead, we concentrated on behavioral and emotional changes in patients who reported primary headaches. Third, we lacked information on patient-level changes in lifestyle habits, such as differences in smartphone usage or exercise routines before and after the emergence of COVID-19.
Future studies should investigate changes in headache characteristics, including frequency and intensity, among this patient population. These changes should be assessed not only before and after the onset of COVID-19 but also during the current period, as the COVID-19 pandemic subsides.
In conclusion, pediatric visitors to our headache clinic exhibited significantly higher levels of anxiety and depression after the onset of the COVID-19 pandemic compared to those who visited in the pre-onset period. As stress-related and psychiatric factors can exacerbate headaches, understanding the profound impact of the pandemic on pediatric headache may provide valuable insights into the management of headaches in children.